

اثرات اسیدآمینه های ال متیونین و ال لیزین بر میزان رشد جمعیت کلنی

هدف از مطالعه حاضر ارزیابی اثرات تغذیه اسیدآمینههای ال متیونین و ال لیزین روی رشد جمعیت کلنی، تخمگذاری ملکه، پرورش نوزادان و میزان تولید عسل بود. این آزمایش از 27 اردیبهشت ماه سال 1398 شروع و به مدت 45 روز تا 10 تیر ماه در استان اردبیل انجام شد. برای این آزمایش تعداد 48 کندو انتخاب شدند و کلنیها از نظر شرایط محیطی، جمعیت، ذخیره گرده و عسل و ملکههای خواهری یکسان سازی شدند.
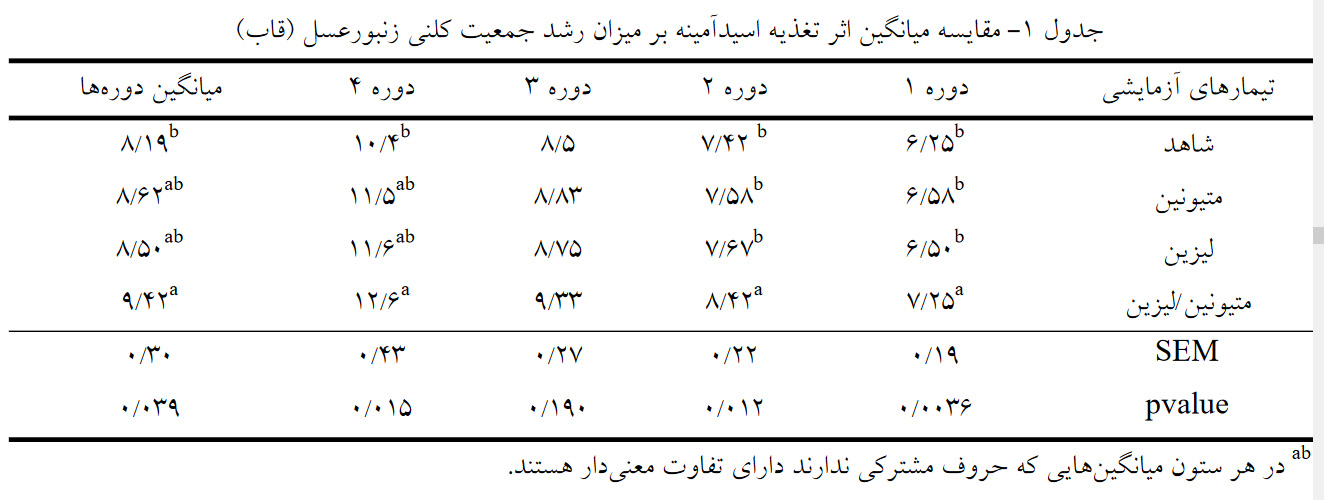
آزمایش در قالب طرح کاملا تصادفی با 4 تیمار و 12 تکرار انجام شد. تیمارها شامل: 1) تیمار شاهد (کلنیهای تغذیه شده با شربت فاقد مکمل اسید آمینه) و تیمارهای 2، 3 و 4 که به ترتیب با شربت مکمل شده با اسید آمینه متیونین، اسید آمینه لیزین و مخلوطی از اسیدآمینههای لیزین و متیونین تغذیه شدند. نتایج نشان داد که در دوره اول و دوم بیشترین میزان جمعیت در تیمار متیونین/لیزین مشاهده شد (05/0 > p). در دوره چهارم و میانگین دورهها، بیشترین جمعیت کلنیها مربوط به تیمار متیونین/لیزین (به ترتیب 6/12 و 42/9 قاب) و کمترین جمعیت مربوط به تیمار شاهد (به ترتیب 4/10 و 19/8 قاب) بود.
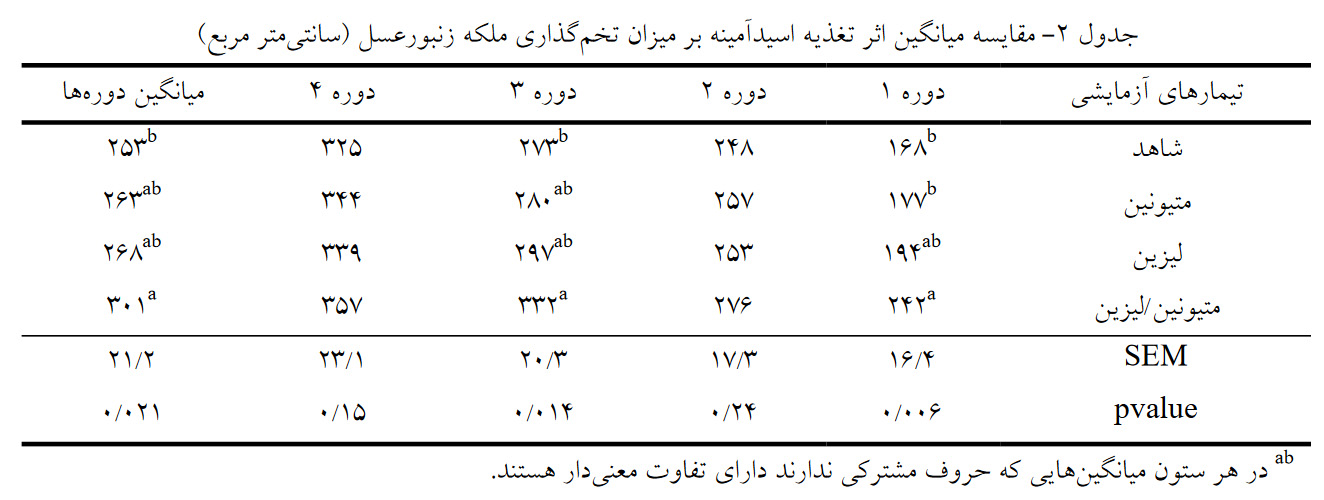
در تمام دورههای آزمایش، تخمگذاری ملکه در زنبورهایی که با مخلوط اسیدآمینههای متیونین/لیزین تغذیه شده بودند در مقایسه با گروه شاهد افزایش معنیداری داشت (05/0 > p). همچنین بهترین عملکرد برای صفت پرورش نوزادان و تولید عسل در تیمار متیونین/لیزین بدست آمد. در کل بررسی ما تاثیر مثبت تغذیه مخلوط اسیدآمینههای لیزین و متیونین بر جمعیت کلنی، تخمریزی ملکه و تولید عسل را تائید میکند.
زنبورعسل اروپایی یا معمولی (Apis mellifera L.) از راسته بال غشاییان و خانواده Apidae است (5). پرورش زنبورعسل رواج گستردهای در جهان داشته و اهمیت اقتصادی آن از ابعاد مختلفی نظیر گردهافشانی و تولید فراوردههایی مانند عسل، موم، بره موم، ژله رویال و زهر که مصارف صنعتی، بهداشتی، غذایی و دارویی دارند به اثبات رسیده است (8).
مواد اصلی در رژیم غذائی کامل زنبورعسل شامل شهد گل و گرده گل میباشند. زنبورهای عسل برای رشد و نمو و فعالیت به کربوهیدراتها، پروتئینها، چربیها، ویتامینها و مواد معدنی نیاز دارند. کربوهیدراتها را بهطور طبیعی از طریق شهد گل و پروتئینها، چربیها ویتامینها و مواد معدنی را از طریق گرده گل دریافت میکنند (7).
حدود 13 درصد از وزن بدن یک زنبور تازه متولد شده و حدود 5/15 درصد از وزن بدن زنبور پنجروزه را مواد پروتئینی تشکیل میدهد (16). هر زنبورعسل از زمان تفریخ تخم تا خارج شدن از سلول، برای رشد خود به 21/3 میلیگرم نیتروژن نیاز دارد، که این مقدار نیتروژن معادل 120 میلیگرم گرده گل میباشد (16). زنبوران عسل از مواد پروتئینی بیشتر جهت تأمین عناصر ساختمانی، ماهیچهها، غدد و سایر بافتها استفاده میکنند.
زنبورهای عسلی که از گرده های گل یا مواد پروتئینی بیشتر تغذیه مینمایند، قادرند تعداد بیشتری از لاروها را تغذیه نمایند (17). رولستون و کان (2000) دامنه تغییرات میزان پروتئین گرده را بسته به نوع گونه بین 5/2 تا 61 درصد متغیر گزارش کردند.

اطلاعات مربوط به پروتئین و دیگر احتیاجات غذایی زنبورعسل نشان میدهد که گردههای حاوی کمتر از 20 درصد پروتئین خام نمیتوانند احتیاجات کلنی را برای تولید مطلوب فراهم سازند (10). زنبورهایی که بیش از 23 درصد پروتئین مصرف میکنند قادرند بهطور موفقیتآمیزی جمعیت نوزادان را افزایش دهند (27). پروتئینها تأمین کننده انواع مختلف آمینواسیدها میباشند. 10 آمینواسید تریونین، والین، متیونین، ایزولوسین، لوسین، فنیل آلانین، هیستیدین، لیزین، آرژنین و تریپتوفان به عنوان آمینواسیدهای ضروری در تغذیه زنبورعسل شناسایی شدهاند (28). اما پروتئین بعضی از گردهها فاقد کلیه اسیدآمینههای مورد نیاز زنبور میباشند (9).
کمبود گرده گل منجر به کاهش میزان تخمگذاری ملکه، کاهش پرورش نوزاد، رشد غیرطبیعی، کاهش طول عمر در کارگران بالغ، کاهش در جمعیت زنبوران نر، افزایش حساسیت به بیماریها، عدم رشد مناسب تخمدانها در ملکه، کاهش تولید اسپرم، تعویض ملکه، جویدن لاروها و شفیرهها، عدم تولید زهر به مقدار کافی، نداشتن توانایی لازم برای زمستان گذرانی، کاهش تولید و یا عدم تولید ژله شاهانه، کاهش تولید موم، کاهش در تولید عسل و حتی منجر به مرگ کلنی میشود (6، 21، 28، 29). در صورتی که گرده گل به مقدار کافی در مزارع موجود نباشد، ضروریست زنبورداران مکمل یا جانشین گرده تازه تهیه نموده و در اختیار کلنیها قرار دهند، و به این صورت رشد و سلامت کلنی را تأمین نمایند (6).
همه حیوانات نیاز به یک منبع غذایی غنی از اسیدهای آمینه ضروری دارند که این اسیدهای آمینه جهت رشد، نگهداری سلولهای بدنی و تولیدمثل استفاده میشوند. اسیدهای آمینه ضروری با مصرف پروتئین موجود در سایر حیوانات و گیاهان به دست میآیند (15و 20). کیفیت و کمیت این آمینواسیدها طول عمر و باروری حشرات را افزایش میدهد (18). شواهد نشان دادهاند که حشرات محلولهای شکر حاوی آمینواسیدها را ترجیح میدهند (22). در مطالعه اینویه و والر (1984) کاهش مصرف شهد توسط زنبورعسل(Apis Mellifera) در زمانی که غلظت آمینواسیدها افزایش مییافت مشاهده شد و فقط در مصرف فنیل آلانین استثنا وجود داشت.
برطبق نتایج کارتر و همکاران (2006) و برتازینی و همکاران (2010)، شهد حاوی اسیدآمینه پرولین بیشتر از شهدهایی که فقط حاوی قند بودند ترجیح داده شد. برطبق نتایج روگالا و سماز (2004) تغذیه ایزومرهای L آمینواسیدها نسبت به ایزومرهای DL نتایج بهتری را نشان دادند. براساس سایر گزارشات ازجمله سامرویل (2005) مطالعه در زمینه تغذیه زنبوران عسل با مواد پروتئینی و اسیدآمینهها و نقش آمینواسیدها در زنبورعسل کمتر صورت گرفته است.
لیزین از اسیدهای آمینه ضروری برای تک معدهایها میباشد که بهاندازه کافی در بدن حیوان ساخته نمیشود و نیاز است که بهصورت سنتتیک به حیوان خورانیده شود. اسیدآمینه لیزین، پایه و اساس ارزیابی دیگر اسیدهای آمینه جیره برای ایجاد تعادل ایدهآل اسیدهای آمینه شناخته شده است. بنابراین مکمل سازی آن در جیره بهمنظور کاهش پروتئین خام جیره امکانپذیر خواهد بود.
لیزین به عنوان اسیدآمینه مرجع در پروتئین بوده و وارد سایر مسیرهای متابولیکی دیگر نمیشود و سطح احتیاجات آن به میزان اندکی توسط سایر فعالیتهای متابولیکی تحت تأثیر قرار میگیرد. متیونین یکی از آمینواسیدهای ضروری در همه سلسلههای بیولوژیکی میباشد که نقش مهمی در متابولیسم اولیه و ثانویه دارد و به عنوان دهنده یک گروه متیل مطرح است. این اسیدآمینه نقش کلیدی در ساخت پروتئین و کاتابولیسم سیستم ایمنی در حیوانات دارد (26).
باتوجه به کمبود گرده در اوایل بهار و نیاز به ایجاد تحریک و فعالیت بیشتر زنبورها و تخمگذاری و همچنین نیاز به وجود اطلاعات بیشتر در خصوص تغذیه با اسیدهای آمینه و تأثیر آن بر زنبورها، هدف از این تحقیق بررسی اثرات دو اسیدآمینه ضروری، دی ال متیونین و ال لیزین بر رشد زنبورعسل و عملکرد آنها مورد بررسی قرارگرفت.
مواد و روشها
این پژوهش از بیست و هفتم اردیبهشت 1398 در زنبورستان تخصصی و سایت الگویی، ترویجی و تحقیقی واقع در استان اردبیل، شهرستان مشکینشهر، روستای میرک شروع و تا 10 تیرماه به مدت 45 روز انجام گردید. دراین آزمایش از تعداد 48 کلنی زنبورعسل ایرانی (Apis Mellifera meda) استفاده شد. بهمنظور به حداقل رساندن اختلافات ژنتیکی بین تیمارهای آزمایشی قبل از انجام عملیات و در اوایل بهار ملکه های خواهری به روش تکثیر مصنوعی (روش پیوند از لاروهای کمتر از 36 ساعت) به تعداد 150 شاخون از یک کندوی آرام و پر محصول تهیه شدند و به کندوچههای پرورش ملکه انتقال یافتند و بعد از جفتگیری به روش طبیعی و بعد از اطمینان از تخمریزی و سلامت ملکه به لحاظ فنوتیپی، به تعداد 48 کندو که دارای جمعیت مناسب بودند معرفی شدند.
جهت تحریک ملکه برای تخمگذاری، تمامی کندوها با شربت متوسط تغذیه شدند. برای تهیه شربت، شکر سفید را در آب ولرم به نسبت 1به1 مخلوط نموده تا کاملاً حل شود. سپس برای هر تیمار 3 لیتر شربت در ظرفهای مخصوص جدا شد. در کنار تغذیه آزاد (پرواز و استفاده از گلهای مزارع) از محلول شربت و اسیدآمینه طبق برنامه و بهصورت روزانه با پیمانهی مدرج به میزان250 میلیلیتر برای هر کندو داده میشد و این کار تا 5 تیرماه تکرار شد.
قبل از شروع آزمایش، کندوها ازنظر جمعیت زنبوران بالغ، تعداد نوزادان، و میزان ذخیره گرده و عسل یکسانسازی شدند. برای اندازهگیری میزان جمعیت از معیار قاب استفاده شد بهطوری که پر بودن دو طرف قاب پوشیده از جمعیت یک قاب محسوب شده و کمتر از آن کسری از عدد یک در نظر گرفته شد. میزان گرده نیز توسط قابهای درجهبندی اندازهگیری شد. به عنوان مثال، میانگین گرده برای کلنیها، یک قاب در نظر گرفته شد و بهمنظور یکسانسازی، کلنیهایی که گرده آنها بیشتر از یک قاب بود از گرده آنها برداشته شد و به کلنیهایی که گرده آنها کمتر از یک قاب بود اضافه شد. بدین ترتیب میزان هرکدام از صفتها در تمامی کلنی ها اندازهگیری شد و سپس براساس میانگین محاسبهشده، تمامی کلنی ها همگن شدند.
بعد از انجام عملیات یکسانسازی و اطمینان از اینکه کندوها دارای جمعیت یکسان و دارای ملکه با توانایی تخمریزی یکسانی هستند برای انجام آزمایش آماده شدند. سپس کندوها بهصورت تصادفی به 4 تیمار (هر تیمار شامل 12 کندوی مدرن لانگستروت) تقسیم شدند. تیمارها شامل: 1) گروه شاهد (شربت شکر روزانه به میزان 250 میلیلیتر بدون اسیدآمینه)، 2) شربت شکر حاوی اسیدآمینه ال متیونین، 3) شربت شکر حاوی اسیدآمینه ال لیزین و 4) شربت شکر حاوی مخلوط اسیدهای آمینه ال متیونین و اللیزین بود.
میزان استفاده از اسیدآمینه ها یک گرم به ازای هر لیتر شربت بود. لازم به ذکر است که اسیدآمینههای مذکور با ترازوی دیجیتال برای مصرف روزانه وزن شده و در جای خشک و خنک و دمای کمتر از 25 درجه سانتیگراد نگهداری میشدند. در این آزمایش از آمینواسید ال متیونین (L-Methionine) با خلوص 99 درصد و با نام اختصاریL-Met100 ساخت کشور مالزی تحت لیسانس کشور کره (سئول) و آمینواسید اللیزین (L-Lysine) با خلوص 99 درصد ساخت کشور اندونزی تحت لیسانس کشور کره (سئول) استفاده شد.
در طول زمان آزمایش هر 11 روز یکبار کندوهای مربوط به هر تیمار مورد بررسی قرارگرفت و میزان جمعیت و میزان تخمگذاری ملکه و پرورش نوزادان (لارو و شفیره) اندازهگیری شدند. برای اندازهگیری میزان جمعیت از معیار قاب استفاده شد، به طوریکه اگر دو طرف قاب پوشیده از جمعیت بودند، یک قاب کامل در نظر گرفته شد و اگر دو طرف قاب بهصورت کامل پوشیده از جمعیت نبود، کسری از عدد یک در نظر گرفته میشد. این صفت در چهار دوره اندازهگیری شد. همچنین میزان تخمگذاری ملکه و پرورش نوزادان (میزان لارو، شفیره) نیز در هر 11روز یکبار اندازهگیری شد.
برای این منظور سطحشان بهوسیله قابهای درجهبندی شده (بر حسب سانتیمتر مربع) اندازه گیری شد. به این صورت که یک کادر خالی بهوسیله سیم به مربعهای 5×5 سانتیمتر مربع تقسیم شد که مساحت هر مربع 25 سانتیمتر مربع بود و در داخل هر مربع 100 سلولشان قرار میگرفت. با قرار دادن قاب خالی روی قابهای حاوی تخم و نوزاد، تعداد مربعها مشخص شد و مساحت آنها محاسبه شدند (10). میزان عسل تولیدی در اواسط مردادماه و 45 روز پس از پایان دوره مصرف اسیدآمینه ها اندازهگیری شد. برای این منظور با وزن کردن قابهای هر کندو، وزن عسل تولیدی در هر تیمار اندازهگیری شد. این مطالعه با 4 تیمار و در 12 تکرار در قالب طرح کاملاً تصادفی انجام شد. دادههای حاصل توسط برنامه SAS 9.2 و با استفاده از رویه GLM مورد تجزیه و تحلیل قرارگرفت. برای مقایسه میانگین تیمارها از آزمون دانکن در سطح 5 درصد استفاده شد.
نتایج
اثر تغذیه اسیدآمینهها بر میزان رشد جمعیت کلنی زنبورعسل: در ابتدا لازم به توضیح است که برای رسیدن به دوزهای اسیدآمینههای مورد استفاده در این آزمایش، یک پیش آزمایشی با آمینواسیدهای ال لیزین و ال متیونین و همچنین ترکیبی از این دو اسیدآمینه در سال 1397 انجام گرفت. به طوریکه کندوها با ترکیب متفاوتی از اسیدآمینهها به میزان 5/0، 1، 5/1و 2 گرم بهصورت مخلوط با یک لیتر شربت یکبهیک تغذیه شدند.
نتایج آزمایش نشان داد که تیمارها تمایلی به مصرف شربتهای حاوی 5/1 و 2 گرم اسیدآمینه را از خودشان نشان ندادند و همچنین مصرف شربت حاوی 5/0 گرم اسیدآمینه تفاوت معنیداری با گروه شاهد در میزان تخمگذاری، جمعیت کلنی و تولید عسل نشان نداد. بنابراین 1 گرم اسیدآمینه در شربت به عنوان بهترین دوز در آزمایش حاضر مورد استفاده قرارگرفت.
نتایج مقایسه میانگین اثر اسیدآمینهها بر میزان جمعیت کلنی در دورههای زمانی مختلف در جدول 1 آورده شده است. نتایج حاصل از تجزیهوتحلیل دادهها نشان داد که در دورههای زمانی 1 و 2 اثر مخلوطی از اسیدآمینههای لیزین و متیونین در تغذیه تکمیلی زنبوران عسل نسبت به گروه شاهد و گروههای لیزین و متیونین معنیدار بود (05/0 > P) و جمعیت کلنی را افزایش داد. اما در دوره سوم آزمایش در این صفت تفاوت معنی داری بین تیمارهای آزمایشی مشاهده نشد (05/0 < P).
جمعیت کلنی ها، در دوره 4 و در میانگین کل دوره ها نیز تفاوت معنیداری بین تیمار چهارم و تیمار شاهد داشت (05/0 > P). به طوریکه تیمار متیونین/لیزین بالاترین (6/12 و 42/9 قاب به ترتیب در دوره 4 و میانگین دورهها) و تیمار شاهد کمترین (4/10 و 19/8 قاب به ترتیب در دوره 4 و میانگین دورهها) میزان جمعیت را داشتند (جدول 1). اما زمانی که اسیدهای آمینه لیزین و متیونین بهصورت جداگانه در تغذیه تکمیلی زنبوران عسل مورد استفاده قرارگرفتند تفاوت معنیداری با سایر تیمارها در دوره 4 و میانگین دورهها نشان ندادند (05/0 > P).
اثر تغذیه اسیدآمینهها بر میزان تخمگذاری ملکه زنبورعسل: همچنانکه در جدول 2 مشاهده میشود در دوره اول بیشترین میزان تخمریزی ملکه در کندوهایی مشاهده شد که با شربت حاوی مخلوطی از اسیدآمینهای لیزین و متیونین تغذیه شده بودند (242 سانتیمتر مربع)، بهطوری که تفاوت این تیمار نسبت به تیمار شاهد و تیمار متیونین (به ترتیب 168 و 177 سانتیمتر مربع) در سطح 5% معنیداری بود. در دوره 3 آزمایش و میانگین کل دورهها نیز بیشترین مقدار تخمریزی ملکه در تیمار متیونین/لیزین مشاهده شد که نسبت به تیمار شاهد ازنظر آماری در سطح 5 درصد معنیدار بود. مصرف اسیدآمینهها بر میزان تخمریزی ملکه در دورههای 2 و 4 تأثیر معنیدار نداشت (05/0< P).
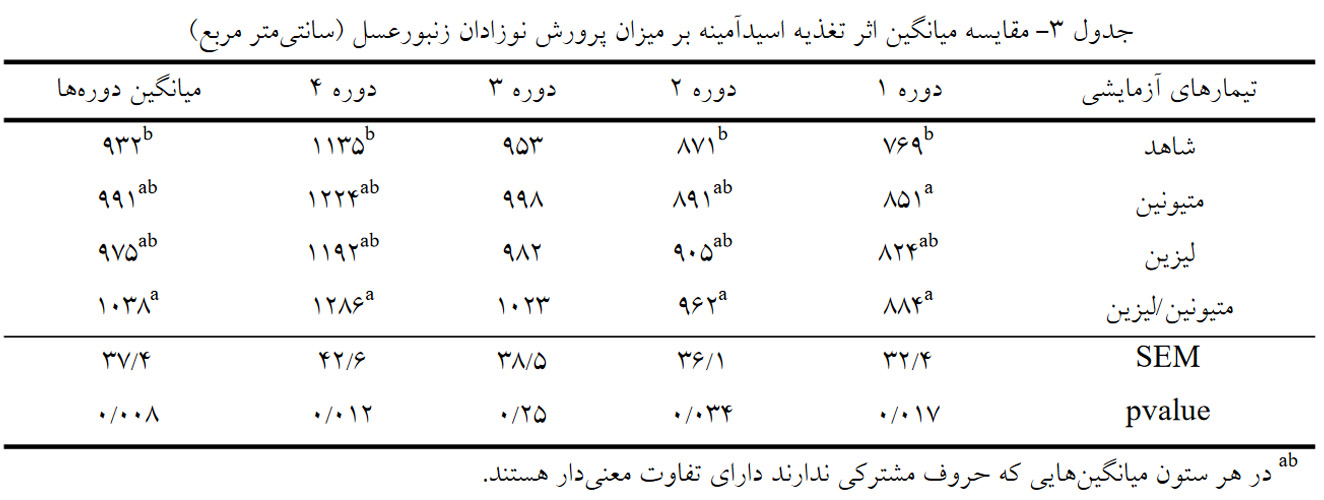
اثر تغذیه اسیدآمینه ها بر میزان پرورش نوزادان زنبورعسل: نتایج اثر اسیدآمینه ها بر میزان پرورش نوزادان در دوره های زمانی مختلف در جدول 3 آورده شده است. نتایج این مطالعه نشان داد که در صفت میزان پرورش نوزادان، تیمار متیونین/لیزین نسبت به تیمار شاهد در دوره های 1، 2، 4 و میانگین دوره ها تأثیر معنی داری داشت (05/0 > P). همچنین در دوره اول، تیمار متیونین نیز میزان پرورش نوزادان را بهطور معنی داری (05/0 > P) نسبت به گروه شاهد افزایش داد (851 در مقابل 769 سانتیمترمربع). میزان پرورش نوزادان در دوره سوم تحت تأثیر مصرف آمینواسیدها قرار نگرفت (05/0< P).
اثر تغذیه اسیدآمینه ها بر میزان تولید عسل: نتایج مقایسه میانگین اثر اسیدآمینه ها بر میزان عسل تولیدی در نمودار 1 آورده شده است. میانگین وزن عسل تولیدی تحت تأثیر مصرف اسیدآمینهها در سطح 05/0 قرارگرفت. به طوری که بیشترین عسل تولیدی از کندوهایی جمعآوری شد که با شربت حاوی مخلوطی از اسیدآمینهای لیزین و متیونین و اسیدآمینه لیزین بهتنهایی، تغذیه شده بودند (به ترتیب 42/11 و 74/10 کیلوگرم). وزن عسل تولیدی در زنبورهایی که با اسیدآمینه متیونین تغذیه شده بودند نسبت به زنبورهایی که با جیره شاهد تغذیه شده بودند، تفاوت معنیداری نشان نداد (05/0< P).
بحث :
در گزارشی امیری و همکاران (1396) سه نوع اسیدآمینه ضروری والین، ایزولوسین و متیونین را به جیره زنبورعسل اضافه نمودند و اثراتشان را روی میزان رشد جمعیت، میزان توسعه تخمریزی و تعداد نوزاد و وزن خشک، پروتئین و چربی لاشه زنبوران عسل مورد بررسی قراردادند.
در این آزمایش آنها دریافتند که استفاده از این اسیدآمینههای ضروری در تغذیه تکمیلی زنبورعسل تأثیری روی میزان رشد جمعیت زنبورعسل در دورههای مختلف ندارد. همچنین آنها نشان دادند که بیشترین میزان تخمگذاری ملکه و پرورش نوزادان در تیمار ایزولوسین و کمترین آن مربوط به تیمار والین بود.
 در این آزمایش بیشترین میزان پروتئین بدن زنبورعسل در تیمار والین مشاهده شد که با تیمار ایزولوسین تفاوتی نداشت اما بهطور معنیداری از میزان پروتئین در تیمارهای شاهد و متیونین بالاتر بود. در تحقیق دگروت (1953) نیز استفاده از اسیدهای آمینه در جیره زنبورعسل، ماده خشک و میزان نیتروژن بدن زنبورهای عسل را افزایش داد و آنها نتیجه گرفتند که استفاده از اسیدآمینه ایزولوسین به میزان 4 درصد، باعث بیشترین افزایش وزن در زنبورعسل میشود. نهضتی (1387) و نیک کار چنیجانی (1392) بیان کردند که هرچقدر جیره غذایی زنبورعسل ازنظر مواد مغذی کاملتر باشد نیتروژن بیشتری در بدن حشره ذخیره میشود.
در این آزمایش بیشترین میزان پروتئین بدن زنبورعسل در تیمار والین مشاهده شد که با تیمار ایزولوسین تفاوتی نداشت اما بهطور معنیداری از میزان پروتئین در تیمارهای شاهد و متیونین بالاتر بود. در تحقیق دگروت (1953) نیز استفاده از اسیدهای آمینه در جیره زنبورعسل، ماده خشک و میزان نیتروژن بدن زنبورهای عسل را افزایش داد و آنها نتیجه گرفتند که استفاده از اسیدآمینه ایزولوسین به میزان 4 درصد، باعث بیشترین افزایش وزن در زنبورعسل میشود. نهضتی (1387) و نیک کار چنیجانی (1392) بیان کردند که هرچقدر جیره غذایی زنبورعسل ازنظر مواد مغذی کاملتر باشد نیتروژن بیشتری در بدن حشره ذخیره میشود.
اسیدهای آمینه میتوانند هم اثرات مهاری و هم اثرات تحریکی روی تغذیه حشرات داشته باشند و گزارش شده است که اثر آمینواسیدها روی عملکرد و رشد حشرات به غلظت اسیدآمینه و نوع گونه وابسته است (12و 13) و نتایج متفاوت در پژوهشهای مختلف در ارتباط با تغذیه اسیدهای آمینه میتواند به همین دلیل باشد. در مطالعهای زمانی که غلظت اسیدهای آمینه (مخلوطی از 24 اسیدآمینه L و 2 اسیدآمینه DL) در جیره افزایش یافت، مصرف شهد توسط زنبوران عسل کاهش یافت (17). در پژوهش دیگری که روی گونههای مختلف زنبورعسل انجام شد، نتیجه گرفتند که افزودن اسیدآمینه به میزان 80-35 میلیمولار بر میزان مصرف شهد زنبوران عسل تأثیر معنیداری ندارد (25).
تعداد مطالعات روی اثرات اسیدآمینهها بر عملکرد زنبوران عسل محدود است اما محققین زیادی اثر منابع مختلف پروتئینی روی عملکرد و شاخصهای رشد زنبوران عسل را مورد بررسی قراردادند. در مطالعهای بابایی و همکاران (1390) گزارش دادند که زنبورانی که با جیرههای حاوی پروتئین بالا تغذیه شده بودند دارای رشد جمعیت بیشتری بودند و سلامت کلنی نیز بهبود یافت.
در آزمایش موسا و همکاران (1989) که آرد سویا و شیر خشک بدون چربی را به عنوان منابع پروتئین در جیره با گرده جایگزین کرده بودند دریافتند که پرورش نوزادان و جمعیت کلنیها در تیمارهای مورد آزمایش بهبود یافت. در پژوهش جواهری و همکاران (1378) که از مکملهای پروتئینی در جیره استفاده شده بود نتیجه گرفتند که استفاده از جانشین گرده و مکمل گرده با پودر کنجاله سویا، پودر گلوتن ذرت و پودر کنجاله کنجد باعث افزایش جمعیت کلنیها و توسعه تخمریزی و تعداد نوزادان میشود.
هرچند که این محققین به دلایلی همچون کم بودن قابلیت هضم این جیرهها و همچنین خوشخوراک نبودن آنها و وجود ترکیبات مضر در این مکملها، مصرف آنها را برای تغذیه تحریکی زنبورها توصیه نکردند. در پژوهش دیگری، مصرف مخلوطی از شکر، گرده و آرد سویا جایگزین گرده در تغذیه تکمیلی زنبورعسل شد که این مخلوط رشد نوزادان را به میزان 81% در زمان کمبود گرده افزایش داد (30). در پژوهشی که از جیرههای مختلف پروتئینی در تغذیه زنبورعسل اروپایی استفاده شده بود، نتیجه گرفتند که کیکهای جانشین گرده و مکملهای گرده باعث افزایش معنیدار در طول عمر زنبوران کارگر، درصد ماده خشک و درصد چربی لاشه زنبوران شده است.
در این آزمایش محققین پیشنهاد دادند که از مواد پروتئینی مانند کنجاله و پودر سویا، مخمر نانوایی و گلوتن گندم میتوان در تهیه مکمل گرده گل و کیکهای جانشین برای مصرف زنبورعسل استفاده کرد (6). رضایی و همکاران (1394) در آزمایشی اثرات مواد پروتئینی تخمیر شده روی زنبورعسل اروپایی را مورد بررسی قراردادند. آنها نتیجه گرفتند زمانی که منابع پروتئینی تخمیر میشوند بر میزان مصرف غذا و میزان توسعه تخمریزی و تعداد نوزادان اثر مثبتی داشته و عملکرد زنبورعسل را افزایش میدهد. در مطالعهای درویش زاده و همکاران (2015) گزارش نموند که افزودن اسیدآمینه پرولین به جیره زنبورعسل، قطر غدههای بالا حلقی و مصرف شربت را افزایش داد.
در تحقیقاتی که روی میزان تمایل زنبوران عسل به انواع اسیدآمینهها انجامشده است، نشان داده شد که تمایل زنبورعسل به مصرف اسیدآمینههای مختلف متفاوت است و به مصرف بعضی از اسیدهای آمینه تمایل بیشتر و به بعضی از آنها تمایل کمتری دارد (11). در آزمایش دیگری، مصرف اسیدهای آمینه فنیل آلانین، هیستیدین، ایزولوسین، لوسین، تریپتوفان، والین و ترئونین در دوره 7 روزه اول تغذیه باعث افزایش وزن خشک زنبورعسل شدند اما در روز 14 در این تیمارها کاهش وزن مشاهده شد (15).
اگرچه اثرات مثبت آمینواسیدها روی شاخصهای فیزیولوژیکی زنبوران عسل در پژوهشهای قبلی گزارش شده است (15)، اما تحقیقات جامعی در ارتباط با اثرات کل اسیدهای آمینه روی میزان تخمگذاری ملکه، توسعه و پرورش نوزادان و میزان تولید عسل وجود ندارد. بنابراین باتوجه به نتایج این آزمایش پیشنهاد میشود اثر دوزهای مختلف اسیدهای آمینه لیزین و متونین روی میزان مصرف خوراک و عملکرد زنبورعسل در دورههای مختلف نیز مورد بررسی قرار گیرد.
نتیجه گیری
بهطور کلی نتایج این پژوهش نشان داد که استفاده از مخلوط اسیدآمینههای لیزین و متیونین در جیره غذایی زنبوران عسل میزان جمعیت کلنی، تخمریزی ملکه و پرورش نوزادان و عسل تولیدی را نسبت به سایر تیمارها بهبود داد. بنابراین با توجه به اثرات مثبت لیزین و متیونین بر عملکرد زنبوران عسل میتوان از مخلوط این اسیدآمینهها در تغذیه تکمیلی زنبوران عسل استفاده نمود و در صنعت زنبورداری بازده اقتصادی را از این طریق افزایش داد.
منابع :
1- امیری، ب.، 1396. بررسی اثر سه اسیدآمینه ضروری والین، متیونین و ایزولوسین بر رشد جمعیت و میزان تخمریزی ملکه زنبورعسل گونه معمولی(Apis mellifera L) ، پایاننامه کارشناسی ارشد، دانشکده علوم دامی و صنایع غذایی، دانشگاه کشاورزی و منابع طبیعی رامین خوزستان.
2- بابایی، س.، نهضتی پاقلعه، غ.، ملک زاده، ح.، و عباسی، س.، 1390. مصرف جیرههای پروبی (Pro Bee) بعنوان مکمل و جانشین پروتئینی در تغذیه کارگران زنبورعسل و تاثیر آن بر وزن، ذخیره پروتئین و چربی( Apis mellifera) . مجله علوم دامی ایران، 33(1)، صفحات 43- 40.
3- جواهری، د.، اسماعیلی، م.، نیکخواه، ع.، میرهادی، ا.، و طهماسبی، غ.، 1378. تغذیه تحریکی زنبوران عسل همراه با مکمل و جانشین پروتئین گرده و اثر آنها در رشد و مقاومت کلنیها و تولید عسل. شرکت جهاد تحقیقات و آموزش. صفحه 76.
4- رضایی، ع.، نهضتی پاقلعه، غ.، مرادی شهربابک، م.، و گنج خانلو، م.، 1394. تأثیر مواد پروتئینی تخمیرشده بر مقدار خوراک مصرفی، پروتئین لاشه، پرورش نوزاد و رشد جمعیت کلنی زنبورعسل اروپایی (Apis mellifera). مجله علوم دامی ایران، 46(3)، صفحات 345- 350.
5- سیاوشی، ی.، بحرینی بهزاد، م.، و بهجتیان اصفهانی، م.، 1396. جداسازی، شناسایی و بررسی تغییرات پروتئینهای غدد شیری زنبور عسل در سنین مختلف رشد به روش الکتروفورز ژل پلی اکریل آمید سدیم دودسیل سولفات، مجله پژوهشهای جانوری ) مجله زیستشناسی ایران)، 30(4)، صفحات 428-435.
6- عباسیان، ع.، و عبادی، ر.، 1381. تأثیر برخی منابع پروتئینی بر طول عمر، پروتئین و چربی بدن زنبورهای کارگر زنبورعسل اروپاییی Apis mellifera L.، علوم و فنون کشاورزی و منابع طبیعی، 6(2)، صفحات 149- 157.
7- محب الدینی، ح.، مقصود لو، ع.، دستار، ب.، و طهماسبی، غ.، 1397. تولید ژله رویال، مقدار تیامین ژله رویال و توسعه غدد هیپوفارنژیال در کلنیهای زنبورعسل ایرانی (Apis mellifera Meda) تغذیه شده با سطوح مختلف تیامین، مجله پژوهشهای جانوری (مجله زیستشناسی ایران)، 31(4)، صفحات 406-417.
8- موسوی، م.، 1389. تعیین تنوع ژنتیکی جمعیت زنبورعسل استانهای گیلان و مازندران با استفاده از نشانگرهای میکروساتلایت، پایاننامه کارشناسی ارشد، گروه علوم دامی، دانشکده کشاورزی، دانشگاه زنجان.
9- نهضتی، غ.، 1387. مطالعه هضم چند مکمل پروتئینی در زنبورعسل. پایاننامه دکتری علوم دامی، دانشکده کشاورزی دانشگاه تهران.
10- نیک کار چنیجانی، م.، 1392. تأثیر سطوح مختلف ویتامین C بر میزان جمعیت، تخمگذاری ملکه و زمستان گذرانی کلنی زنبورعسل Apis mellifera، پایاننامه کارشناسی ارشد، گروه علوم دامی، دانشکده کشاورزی، دانشگاه آزاد اسلامی واحد رشت.
11- Bertazzini, M., Medrzycki, P., Bortolotti, L., Maistrello, L., and Forlani, G., 2010. Amino acid content and nectar choice by forager honeybees (Apis mellifera L.), Amino Acids, 39(1), PP: 315-318.
12- Carter, C., Shafir, S. H., Yehonatan, L., Palmer, R. G., and Thornburg, R., 2006. A Novel Role For Prolin Inplant Floral Nectars, Naturwissenschaften, 93, PP: 72-79.
13- Chang, C. H. L., 2004. Effect of Amino Acids on Larvae and Adults of Ceratitis Capitata (Diptera: Tephritidae), Annals of The Entomological Society of America, 97(3), PP: 529-535.
14- Darvishzadeh, A., Hosseininaveh, V., Nehzati, G. H., and Nozari, J., 2015. Effect of Prolin As A Nutrient on Hypopharyngeal Glands During Development of Apis Mellifera (Hymenoptera: Apidae). Arthropods, 4(4), PP: 137-143.
15- De Groot, A. P., 1953. Protein and Amino Acid Requirements of The Honeybee (Apis Mellifica L.). Phys Comp Oec, 3, PP: 197-285.
16- Haydak, M. H., 1970. Honey Bee Nutrition. Annual Review Entomol, 15, PP: 143-156.
17- Inouye, D. W., and Waller, G. D., 1984. Responses Of Honey Bees (Apis Mellifera) To Amino Acid Solutions Mimicking Floral Nectars. Ecology, 65, PP: 618-625 (Abstract).
18- Mevi-Schutz, J., and Erhardt, A., 2005. Amino Acids in Nectar Enhance Butterfly Fecundity: A Long-Awaited Link, The American Naturalist, 165(4), PP: 411-420.
19- Musa, F. H. A., Abdalla, M. R., and E-Sarrag, M. S. A., 1989. Studies on Feeding Colonies Of Honey Bees In Sudan, In: Proceedings of the 4th international conference on apiculture in tropical climates, Cairo, PP: 27-28.
20- Paoli, P. P., Donley, D., Stabler, D., Saseendranth, A., Nicolson, S. W., Simpson, S. J., and Wright, G. A., 2014. Nutritional Balance of Essential Amino Acids and Carbohydrates of The Adult Worker Honeybee Depends on Age, Amino Acids, 46, PP: 1449-1458.
21- Rammesh, R., and Pankiw, T., 2007. Effects of Protein- Constraind Drood Food On Honey Bee (Apis Mellifera L.) Pollen Foragin And Colony Growth. British Ecological Society, 61, PP: 1471-1478.
22- Rathman, E. S., Lanza, J., and Wilsan, J., 1990. Feeding Preferences of Flesh Flies Sarcophaga Bullata For Sugar-Only Vs Sugar-Amino Acid Nectars, American Midland Naturalist, 1242, PP: 379-389 (Abstract).
23- Rogala, R., and Szymas, B., 2004. Nutritional Value for Bees of Pollen Substitute Enriched with Synthetic Amino Acids Part II, Biological Methods, Journal of Apicultural Science, 48(1), PP: 29-36.
24- Roulston, T. H., and Cane, J. H., 2000. Pollen nutritional content and digestibility for animals, Plant Systematics and Evolution, 222, PP: 187-209.
25- Roubik, D. W., Yanega, D., Aluja S. M., Buchmann, S. L., and Inouye, D. W., 1995. On Optimal Nectar Foraging by Some Tropical Bees (Hymenoptera: Apidae), Springer Verlag, 26(3), PP: 197-211.
26- Ruan, T., Li, L., Peng, X., and Wu, B., 2017. Effects of methionine on the immune function in animals, Health, 9, PP: 857-869.
27- Somerville, D., and Nicol, H. I., 2002. Mineral Content of Honeybee-Collected Pollen from Southern New South Wales, Australian Journal of Experimental Agriculture, 42, PP: 1131-1136.
28- Somerville, D., 2005. Fat Bees, Skinny Bees, A Manual On Honey Bee Nutrition for Beekeepers, RIRDC Publication, No 05/054.
29- Szymas, B., and Jedruszuk, A., 2003. The Influence of Different Diets on Haemocytes of Adult Worker Honey Bees, Apis Mellifera, Apidologie, 34, PP: 97–102.
30- Wali-Ur, R., 1991. Management, Studies to Overcome Adversities in Bee Culture, Pakistan Journal of Forestry, 41(3), PP: 130-134.


